
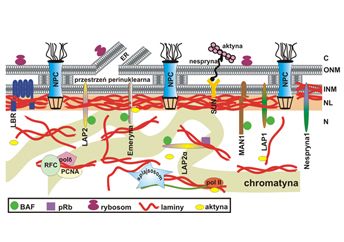
TYPY LAMIN
Laminy są głównym składnikiem blaszki jądrowej - struktury wyścielającej nukleoplazmatyczną powierzchnię wewnętrznej błony jądrowej i stanowiącej element otoczki jądrowej. Obok lokalizacji w otoczce jądrowej, laminy wykrywane są także wewnątrz jądra, w nukleoplazmie. Nie wiadomo, czy występują tam jako dimery, tetramery lub oligomery, czy może formy wysokospolimeryzowane. Laminy identyfikowano u wielokomórkowych eukariontów poza roślinami i grzybami, nie potwierdzono ich obecności w badanych jednokomórkowych organizmach eukariotycznych. Ze względu na wzór ekspresji, sekwencję aminokwasową, właściwości biochemiczne i lokalizację wewnątrzkomórkową laminy można podzielić na dwie główne klasy: laminy A/C oraz laminy B. Laminy typu B występują we wszystkich rodzajach komórek organizmu, w czasie mitozy w większości pozostają połączone z frakcją błonową (pęcherzykami mitotycznymi), mają punkt izoelektryczny w pH<7, są permanentnie izoprenylowane. Z kolei, ekspresja lamin typu A/C jest zależna od stadium rozwoju, identyfikowane są one w komórkach różnicujących się lub zróżnicowanych, podczas podziału jądra komórkowego występują w formie rozpuszczalnej lokalizując się w cytoplazmie, mają charakter zasadowy (zasadowy punkt izoelektryczny) [Rzepecki, 2002 http://www.ncbi.nlm.nih.gov/pubmed/12511969].
U człowieka występują trzy geny lamin: LMNA (zlokalizowany na chromosomie 1q21.2-q21.3), LMNB1 (zlokalizowany na chromosomie 5q23.3-q31.1) oraz LMNB2 (zlokalizowany na chromosomie 19q13.3), które poprzez proces alternatywnego dojrzewania mRNA i potranslacyjnej modyfikacji są podstawą do syntezy 7 izoform białkowych. Laminy A, C, C2, i AΔ10 są syntetyzowane na bazie genu LMNA, który składa się z 12 eksonów. Lamina C kodowana jest przez eksony 1-9 oraz ekson 10. Lamina A powstaje w procesie różnicowego cięcia i składania mRNA (alternatywnego splajsingu), poprzez dodanie eksonu 11 i 12 oraz usunięcie części eksonu 10. W rezultacie powstaje pre-lamina A, która aby mogła zostać zakotwiczona do błony jądrowej, wymaga potranslacyjnej obróbki W efekcie lamina A i lamina C są identyczne w sekwencji początkowych 566 aminokwasów i różnią się tylko fragmentem C-końcowym.
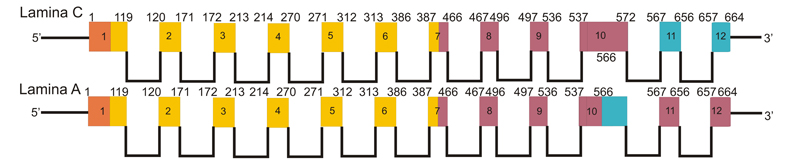
Schemat struktury mRNA laminy C i laminy A
(produkty alternatywnego splajsingu genu LMNA)Gen LMNA zawiera 12 eksonów. Lamina C kodowana jest przez eksony 1-9 oraz ekson 10. Lamina A powstaje w wyniku alternatywnego splajsingu, poprzez dodanie eksonu 11 i 12 oraz usunięcie części eksonu 10. Liczby powyżej schematów wskazują reszty aminokwasowe w powstającym białku, kolory odpowiadają domenom białka (pomarańczowy- domena głowowa, żółty-domena centralna, fioletowy- domena C-końcowa).
Lamina C2 ekspresjonowana jest w komórkach spermatocytów, nie jest ona równomiernie rozlokowana w strukturach otoczki jądrowej, lecz tworzy swego rodzaju domeny. Występująca w komórkach nowotworowych oraz prawidłowych komórkach somatycznych lamina AΔ10 charakteryzuje się skróceniem białka o 30 aa względem laminy A (w czasie obróbki usuwany jest 90 nukleotydowy fragment eksonu 10). Lamina B1 jest jedynym produktem genu LMNB1, z kolei, gen LMNB2 koduje laminy B2 i B3 [Machiels i wsp., 1996 http://www.ncbi.nlm.nih.gov/pubmed/8621584].
Projekty i aktualności:
Projekt finansowany ze środków Wrocławskiego Centrum Badań EIT+ w ramach realizacji projektu 'Biotechnologia i zaawansowane technologie medyczne' - BioMed (POIG.01.01.02-02-003/08) finansowanego ze środków Europejskiego Funduszu Rozwoju Regionalnego (Program Operacyjny Innowacyjna Gospodarka, Poddziałanie 1.1.2).
